Within current evolutionary theory, the primary question to ask about any feature of a living organism is, What adaptive function does it serve? What competitive advantage does it give the organism struggling to survive and reproduce?
The question is legitimate as far as it goes - which isn't very far. No one's going to quarrel with the fact that every organism found in the fossil record had to have been capable of surviving until it went extinct — that is, until it was no longer capable of surviving. (For some fascinating, non-trivial discussion relating to this trivial point, see Ronald Brady's "Dogma and Doubt.") And it is certainly worth investigating how the various features of an organism help to make it fit for the requirements of its life within a particular environment. That's part of getting to know what sort of organism (and environment) we're dealing with.
But when this investigation is narrowed down to a search for survival mechanisms offering a competitive advantage - when the explanatory significance of every feature is reduced finally to the terms of a quantitative judgment about fitness to survive, so that the feature itself is not taken to express anything significant apart from its contribution to survival - when the artful pattern on the butterfly's wing becomes no more than, say, a deceit aimed at birds to avoid being eaten - then we lose the organism as such. We lose it because we're not really seeing at it; we're not allowing its features to speak for themselves, in their own terms. Everything has to be reduced to fit an interpretation that says it is for something else.
Do we actually find in any organism that its life coheres solely around its ability to remain present in its particular environment at the end of the day? Clearly, there are countless different ways organisms can live in the world. All these ways meet the requirements of survivability and of consistency with the laws of chemistry and physics. Is all the rest, then — the expressive unity of an eagle or octopus, the radically differing aesthetic coherence of willow and redwood — rooted in chance? Do we find that there are no lawful patterns expressed in the way evolving species pick one path rather than another through the infinite landscape of survival possibilities? And if there are such patterns, can an evolutionary theory that has nothing to say about them be at all adequate as an explanation of the organism?
Naturalists may develop a profound sense for the inherent lawfulness of a particular organism's way of being — or, we could just as well say, for the organism as a being. But unfortunately naturalists do not have much standing in the age of molecular biology. If, however, it is true that the organism is a recognizable unity; if there are consistent harmonies sounding through its various "survival mechanisms," bringing them together in one song - a song as distinct from those of other organisms as a lullaby is from a dirge; if the organism, not only as a product but also as a shaper of its environment, takes up its creative opportunities and employs them with all the coherence and expressive focus we find in the work of a perceptive artist; if, in sum, there shines consistently through all the morphological, physiological, and behavioral details of an organism a character declaring something more than "I have survived," but also "I am my own sort of being, unified, bound by a lawfulness not only of matter but also of form, and this lawfulness is accessible to those who approach me respectfully" - well, then, the supposedly random foundations underlying contemporary evolutionary theory will have crumbled beneath us.
And, apart from the individual species, it is possible that life on a grander scale may exhibit lawful harmonies. What if we find a certain lawfulness of form and pattern expressed across many different, distantly related groups of organisms occupying radically different environments? It happens that two papers, one dealing with the plumage patterns of birds, and one surveying the "morphodynamics" of dinosaurs, have recently been published by scientists working out of a phenomenological tradition. Both explore patterns of significant form that stretch functional (adaptive) explanations beyond the breaking point.
Dinosaur Morphodynamics
Martin Lockley, a leading dinosaur expert, is curator of the world's largest fossil footprint collection, housed at the University of Colorado in Denver. Some years ago he encountered Wolfgang Schad's work on the form of mammals (1977) and began wondering about its applicability to dinosaurs. Since then a great deal of research has convinced him that the relevance of Schad's approach to his own studies is indeed profound, and this has led to a number of provocative publications exploring what he calls the "morphodynamics" of dinosaurs.
Schad traced the play of certain polarities revealed in the behavior and form of mammals. In particular, there is the polarity between the "nerve-sense" pole on the one hand and the "metabolic" pole at the other. Rodents - nervous, frenetic, alert, and sensitive to their surroundings - live more strongly in their nerves and senses, while the large ungulates (for example, the cow, with its powerful digestive processes and rather more sleepy, "ruminating" relation to its environment) tend toward the metabolic pole. Mediating the two poles and helping to balance them are the rhythmic functions of breathing and blood circulation, which achieve something like their fullest balance in the carnivores such as lions and wolves.
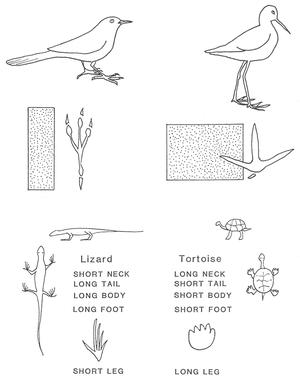
Figure 1. The expression of opposing tendencies. A typical songbird (top left) is smaller, with a relatively narrow, elongated body and a posterior emphasis: long tail, head not distinct from the rest of the body, short bill. The wading shorebird is larger, with a wider body and anterior emphasis: long neck and bill, head distinct from the rest of the body, short tail. The same polar contrast can be seen in the footprints of the birds. And similarly coordinated contrasts are evident in the lizard and tortoise. (Illustration courtesy of Martin Lockley.)Any attempt to characterize in a brief space the way this polarity is expressed in the animal kingdom almost inevitably becomes schematic and misleading; the faithful grasp of polarity requires a lively imagination immersed in the rich world of phenomena. And so we will make no attempt to summarize Schad's valuable findings here, instead referring the reader to his masterwork, Man and Mammals. (Fortunately, this difficult-to-obtain book is now being revised for a new edition.)
However, two of Schad's particular observations need mentioning in the present context. One is that organisms whose inward, physiological orientation is toward the nerves and senses show an outward accentuation of the body's posterior region: for example, the head is not dramatically separated from the rest of the body (think of the mouse), a long tail may be present, and the hind limbs tend to be stronger than the more delicately formed forelimbs, which may be more adapted for handling and sensing things. Contrariwise, organisms with a strong metabolic orientation tend toward an anterior accentuation (here you can think of the bison or giraffe, both with reduced hindquarters and dramatic emphasis on the head region).
The second important point is that the polar dynamic works not only across the animal kingdom viewed as a whole, but also within each group of animals. So it is that among the ungulates, with their strongly developed digestive systems, the bison exemplifies the least development of the nerve-sense pole, while the giraffe, with its more refined head raised high above its digestive organs and alert to the larger environment, shows a relatively strong nerve-sense emphasis. And just as the lion and wolf occupy a middle place within the animal kingdom as a whole, the deer, with its nerve-sense and digestive functions more or less in balance, holds a middle ground among the ungulates.
It's also worth mentioning that the polarity we are speaking about here bears on evolution as well. The general pattern within any group shows an evolution from smaller, more active, nerve-sense-dominated animal forms toward larger, less active, metabolically oriented forms. The latter represent a kind of conclusion, after which evolution takes up a fresh start from another point, just as, upon the extinction of the dinosaurs, the tiny mammalian forms existent at the time became the basis for a new evolutionary thrust.
All of which brings us back to Lockley's work with dinosaurs (2007). He writes that, within the two main dinosaur groups,
one can detect a spectrum of form between small, long-tailed, narrow-bodied, bipedal species (posterior emphasis) and large, wide-bodied, shorter-tailed, quadrupedal species with various cranial processes (crests and horns), indicating an anterior emphasis. These same or similar formative movements reiterate at many different taxonomic levels, and even reiterate within organs of the whole organism such as skulls and feet. (p. 48)Noting that these morphodynamics of dinosaurs can also be recognized among birds and ungulates, Lockley recommends that paleontologists pay much more attention to such patterns of form, and he suggests that the "traditional emphasis on Darwinian functionalism will assume less importance, while the significance of inherent morphodynamics becomes more fully appreciated" (p. 48). After all, we can't be so easily satisfied with the explanation that some particular dinosaur developed a large head "for use in combat" when we find that the movement toward larger heads happens repeatedly and lawfully - in harmonious relation to many other morphological trends - within every group of dinosaurs.
This, of course, amounts to a startling rejection of conventional evolutionary reasoning. The typical causal, deterministic language of biology is simply ill-suited to an understanding of changing patterns of form. Lockley formulates this rule: "rather than single organs changing for specific adaptive purposes, all organs may change simultaneously as part of a shift in a complex, highly dynamic organic system." And what is true of the individual organism seems to be true also of the way species are ordered within higher taxonomic groups, so that "the evolution of species may be, at least in part, an inherent biological dynamic associated with large-scale evolutionary shifts affecting multiple species" (p. 47, emphasis in original).

Figure 2. Evolution within the cerapodan group of dinosaurs. A smaller, ancestral form, along with its footprint, is shown at left, with successive later forms to the right. Relationships similar to those illustrated in Figure 1 are evident and can also be correlated with behavioral patterns, so far as these are known. (Illustration courtesy of Martin Lockley.)This dramatic claim leads to another one of those bombshells Lockley rather casually drops throughout his paper as he unfolds the implications of wide-ranging, repeated patterns of form in the animal kingdom. He notes that different animal groups show three sequential cycles, first of posterior, then of balanced, and then of anterior development, and that the great dinosaur extinction terminating the Mesozoic era came at "precisely the end" of the third cycle, "when horned dinosaurs (like Triceratops) had developed maximum head size.... This seems to be a rather remarkable coincidence, in which a large-scale, inherent biological cycle coincides so precisely with a purported extrinsic cause (meteorite or comet impact)." He goes on:
If a significant number of morphodynamic cycles, culminating in anterior (metabolic) specialization, also result in, or coincide with, extinction, the implication is that extinction, at least to some degree, is an inherent, biological dynamic analogous to a large-scale "life cycle".... Therefore, efforts to seek external causes may be unnecessary and result in misleading, or at best incomplete, explanations and correlations. (pp. 47-8)And in yet another jettisoning of standard evolutionary thought, Lockley questions whether evolution proceeds "by some process of random mutation." After all, given repeated and dynamic morphological tendencies, exhibited widely among different animal groups and manifesting their own relational lawfulness, it is hard to reconcile these with purely random processes. If, as Lockley suggests, "it may be possible to predict the general form and physiology of the whole animal from an analysis or understanding of the parts" (p. 27), and if a similar coherence of form exists within the "superorganisms" comprising the various taxonomic groups, then we are a long way from both the usual adaptationist explanations of the features of animals, and also from chance as the primary generator of variation for natural selection to act upon.
More generally, Lockley argues for a holistic approach to animal morphology, rather than an attempt at feature-by-feature explanation. The latter focuses upon adaptive function (horns are used for butting) whereas the former reckons with the fact that in any organism the modification of one part "will lead to a compensation or ripple effect throughout the whole" organism (p. 27). Lockley quotes the 19th century student of animal form, Geoffrey Saint-Hilaire, who remarked about the unity of the animal kingdom that "There is only a single animal modified by the inverse reciprocal variation of some or all of its parts" (p. 31).
Expressive Feathers
Mark Riegner, who teaches in the Environmental Studies Program at Prescott College in Arizona, has long been interested in animal form, as indicated by the title to his book chapter, "Horns, Hooves, Spots, and Stripes: Form and Pattern in Mammals" (Riegner 1998). He, too, acknowledges a debt to Schad's work, and has tried to extend the Schadian approach to a consideration of animal color patterns, among other things. His most recent paper, published in The Condor (2008), is entitled, "Parallel Evolution of Plumage Pattern and Coloration in Birds: Implications for Defining Avian Morphospace." In this paper he reports the results of a survey of plumage patterns in over 8300 bird species representing 180 different families.
As Riegner notes, plumage patterns and colors have puzzled biologists at least since Darwin's time. Darwin himself proposed that, in bird species where the male is visually striking, this results from females selecting mates possessing the more pronounced features. Others, Riegner writes, have proposed "adaptive hypotheses that assert a functional role of plumage coloration in predator avoidance, social signaling, thermoregulation, abrasion reduction, and foraging efficiency", none of which have proved convincing to the majority of biologists. "Furthermore, it is unclear why similar plumage patterns repeatedly evolve in distantly related lineages or why strikingly different plumage configurations may be exhibited in closely related taxa [taxonomic groups]" (p. 600).
Riegner compared plumage patterns to bird length and bird mass. The plumage patterns he considered were roughly classified as follows: (1) countershaded (for example, brown or gray on the upper, or dorsal, part of the body, white or light-colored on the under, or ventral, part); (2) streaked (ventral longitudinal stripes); (3) blended (for example, uniform gray), spotted, or drab; (4) barred (ventral transverse stripes); (5) bold separation of black and white, or else uniform dark; and (6) reverse countershaded.
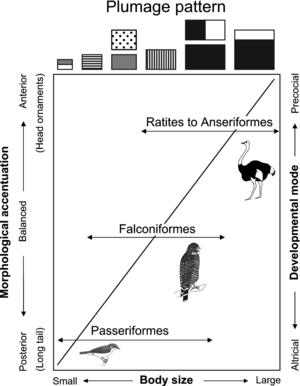
Figure 3. Trajectory of avian plumage patterns and associated morphological features. The same picture recurs at finer levels of analysis; that is, one finds a similar trajectory if one looks only at the passeriforms (generally: perching songbirds) or at various other groups. The plumage patterns shown in the rectangles at the top are (left to right): countershaded; streaked; spotted, drab, or blended; barred; bold separation of black and white, or uniform dark; and reverse countershaded. (Illustration courtesy of Mark Riegner.) What he found, then, was a "trajectory of avian plumage patterns and associated morphological features," as summarized in Figure 3. Here he correlates different plumage patterns not only with body size, but also with the tendency for either the anterior or posterior pole to be emphasized, and with the relative dependence or independence of newborn birds. ("Altricial" indicates that the hatchlings are featherless, blind, and dependent on their parents for food. "Precocial" hatchlings are down-covered, open-eyed, and ready to leave the nest within one or a few days.)
A complicating, and highly significant, observation is that the trajectory is repeated within the individual subgroups constituting the larger pattern. And so there is the same tendency for a shift in plumage pattern as you move from the smaller to larger species within the passeriform subgroup (consisting mostly of altricial songbirds with perching habits) as there is within the avian group as a whole.
Figure 4. The black-and-white warbler (top left), song sparrow (top right), and brown creeper (center left) exhibit plumage patterns typical of small-bodied songbirds, that is, countershading in all three with superimposed streaks in the first two species. The red-bellied woodpecker (center right) and hairy woodpecker (bottom left) are relatively small-bodied woodpeckers compared to the pileated woodpecker (bottom right). Accordingly, the first two have a small head and a countershaded plumage whereas the latter shows an anterior accentuation in the large head with conspicuous crest, as well as dark undersides. (Photos courtesy of John McKean.) 
Such patterns, according to Riegner, "raise questions concerning the validity of nondirectional variation, based on random mutation, as the generator of morphological features" and they "downplay functional mechanisms as the generators of color pattern and biological form." He cites one investigator (C. L. Booth) who has even suggested that "a colouration pattern may have no adaptive significance whatsoever." In any case, he adds, "functional explanations alone are insufficient to understand just why certain chromatic and morphological features covary so consistently" across distantly related groups - that is, why smaller birds tend toward one end of the plumage pattern trajectory, while larger birds tend toward the other end (p. 611).
While the overall trajectory Riegner presents is highly significant statistically, he notes that there are numerous exceptions. Many interacting factors are doubtless at play in the coloration and patterning of bird plumage, and there is no intent here to capture the phenomena within a fixed scheme. However, the statistically significant correlation between plumage patterns and other morphological features of birds across widely separated taxa inhabiting diverse habitats strongly suggests that functional explanations are not by themselves adequate to explain the facts.


Figure 5. Two examples of birds at the upper right portion of the plumage trajectory (Figure 3) for their particular groups. The white-necked raven (left), one of the largest members of the genus Corvus, shows an anterior accentuation with its massive bill, together with a white patch on the upper part of its body (reverse countershading). The large-bodied magpie goose (right) also exhibits anterior emphasis (the unusual peaked forehead), as well as separation of bold black and white plumage patches. The yellow-billed loon (not shown) also has a peaked forehead; it is the largest species of loon. (Raven photo courtesy of Greg Meyer; goose photo by Mark Riegner.)
Toward a More Living Understanding of Evolution
Carl Safina, a MacArthur Fellow and president of the Blue Ocean Institute, recently wrote an essay for the New York Times entitled "Darwinism Must Die So That Evolution May Live." His point was that
evolution can seem uniquely stuck on its founder. We don't call astronomy Copernicanism, nor gravity Newtonism. "Darwinism" implies an ideology adhering to one man's dictates, like Marxism. And "isms" (capitalism, Catholicism, racism) are not science. "Darwinism" implies that biological scientists "believe in" Darwin's "theory." It's as if, since 1860, scientists have just ditto-headed Darwin rather than challenging and testing his ideas, or adding vast new knowledge. (Safina 2009)It is indeed odd that, 150 years after the publication of Origin of Species, the contemporary biological fraternity still expects a kind of ritual declaration of faith in one man and his particular formulation of evolutionary theory, and that those who fail this creedal test are immediately suspected of being creationists or, more generally, "unbelievers." There seems to be a lot more at work here than objective, free-spirited scientific investigation. It's as if the possibility that we might discover meaning and significance, order and pattern - anything more than chance - in the world's living forms touches some deep place of insecurity in the keepers of evolutionary orthodoxy.
We can hope, then, that papers such as the two cited above will play their part in freeing the scientific spirit for a wider attention to the wonders of life on earth.
References
Lockley, Martin G. (2007). "The Morphodynamics of Dinosaurs, Other Archosaurs, and Their Trackways: Holistic Insights into Relationships between Feet, Limbs, and the Whole Body," SEPM Special Publication no. 88 (Society for Sedimentary Geology), pp. 27- 51.
Riegner, Mark (1998). "Horns, Hooves, Spots, and Stripes: Form and Pattern in Mammals," in Goethe's Way of Science: A Phenomenology of Nature, edited by David Seamon and Arthur Zajonc. Albany NY: SUNY Press, pp. 177-212.
Riegner, Mark (2008). "Parallel Evolution of Plumage Pattern and Coloration in Birds: Implications for Defining Avian Morphospace," The Condor vol. 110, no. 4, pp. 599-614.
Safina, Carl (2009). "Darwinism Must Die So That Evolution May Live," New York Times (Feb. 9). Available online: http://www.nytimes.com/2009/02/10/science/10essa.html.
Schad, Wolfgang (1977). Man and Mammals: Toward a Biology of Form, translated by Carroll Scherer. Garden City NY: Waldorf Press.
ABOUT THIS NEWSLETTER
NetFuture, a freely distributed electronic newsletter, is published and copyrighted by The Nature Institute. The editor is Steve Talbott, author of Devices of the Soul: Battling for Our Selves in the Age of Machines (http://natureinstitute.org/txt/st). You may redistribute this newsletter for noncommercial purposes. You may also redistribute individual articles in their entirety, provided the NetFuture url and this paragraph are attached.
NetFuture is supported by freely given reader contributions, and could not survive without them. For details and special offers, see http://netfuture.org/support.html .
Current and past issues of NetFuture are available on the Web:
http://netfuture.orgTo subscribe or unsubscribe, go to
http://netfuture.org/subscribe.html.If you have problems subscribing or unsubscribing, send mail to: [email protected].





This issue of NetFuture: http://netfuture.org/2009/Oct2109_178.html.
